Процесс инкубации яиц (четвёртая часть).
Периоды развития зародыша курицы (часть вторая)
Переломным моментом в жизни зародыша является 15-й день инкубации. В это время резко увеличивается использование веществ желтка, основным источником энергии становится жир. Используется и окисляется большое количество питательных веществ; освобождается большое количество тепла, температура внутри яйца быстро возрастает; появляются первые признаки развивающейся химической терморегуляции. Усиление отдачи тепла яйцом вызывает повышение использования белка и затем желтка; рост зародыша ускоряется. Задержка выделения тепла из яйца приводит к уменьшению использования белка и желтка, рост зародыша замедляется.
Испарение воды из яйца (из аллантоиса) значительно увеличивается. При этом внешние условия влияют на испарение воды из яйца косвенно, изменяя обмен веществ: испарение воды увеличивается вместе с усилением обмена веществ. Для этого периода характерно полное использование белка и втягивание желточного мешка в полость тела зародыша.
Период последних дней инкубации (20-21) отличается от предыдущих. В этот период увеличивается поглощение кислорода и выделение углекислоты, в воздухе после проклева скорлупы появляется аммиак;поглощаются большие количества питательных веществ, образуется большое количество тепла, до наклева резко повышается температура яйца. После проклева быстро испаряются остатки околоплодных жидкостей, тело зародыша сильно охлаждается.
Описанная периодизация зародышевого развития курицы была положена в основу разработки метода дифференцирования режима инкубирования в инкубаторах различных типов. Ее также использовали и при разработке приемов биологического контроля за развитием зародышей.
В качестве основы деления зародышевого развития на периоды чаще принимают морфологические изменения. Г. А. Шмидт устанавливает стадии по изменениям аппаратов снабжения развивающегося внутрилицевой скорлупы организма пищей и кислородом. Он различает соответствующие этим стадиям периоды морфологических, формообразовательных изменений зародыша.
Г. А. Шмидт различает у цыпленка 6 таких стадий развития:
I - стадия осмотического питания и дыхания, в основном за счет кислорода пищи - от яйцекладки до тридцати часов насиживания.
II - стадия получения кислорода и пищи посредством кровеносных сосудов желточного мешка - от 30 часов насиживания до 6 суток;
III - стадия получения кислорода через аллантоис и желточный мешок и пищи из желточного мешка - с 6 по 10-е сутки насиживания;
IV - стадия получения кислорода посредством аллантоиса и пищи через желточный мешок и аллантоис - с 10 по 16-е сутки насиживания;
V - стадия получения кислорода через аллантоис и пищи из желточного мешка - с 16 по 19-е сутки насиживания;
VI - стадия получения кислорода из воздуха и пищи из желточного мешка - 20-21-й день насиживания. Эти периоды деления зародышевого развития курицы довольно полно совпадают с периодизацией, ранее установленной на иной основе М. В. Орловым.
Связь развивающегося зародыша птицы с окружающей средой Г. А. Шмидт видит в развитии эмбриональных приспособлений к обмену веществ, которые обусловливают формообразовательные процессы самого зародыша. Последний вместе с оболочками как бы выделяется из яйца. При этом состояние и свойства белка, желтка и скорлупы, зависящие не только от условий, в которых они формировались в материнском организме, но и от внешних для яйца условий инкубатора, во внимание приняты не были.
В своих последующих работах Г. А. Шмидт устанавливает уже иные стадии развития зародыша курицы. Несколько раньше Г. А. Шмидт выделил как 1-ю стадию период развития в яйцеводе, характеризующуюся внутриклеточным питанием и частично также дыханием. II и III стадии совпадают с I и II стадиями первоначального деления. Но развитие с 6-го дня подразделяется уже не на четыре, как было первоначально, а на три стадии. IV стадия характеризуется тем, что пища доставляется зародышу из желточного мешка, а кислород начинает поступать через сосуды аллантоиса; эта стадия длится с 6 по 12-е сутки; V стадия, во время которой аллантоис сохраняет значение зародышевого органа дыхания и мочевого мешка, а, помимо желтка, развивающийся цыпленок заглатывает и переваривает внутри кишечника белковую массу, длится с 13 по 18-е сутки инкубации; VI стадия - начало дыхания цыпленка (близкого к вылуплению) атмосферным воздухом и питания за счет остатков желтка в желточном мешке, который переходит в брюшную полость цыпленка.С тех же позиций подошли к разработке стадийного анализа разви* тия зародыша птиц М. Д. Попов и Н. П. Третьяков. Эти авторы предлагают вначале различать четыре периода, во время которых наблюдаются сдвиги, направленные к усложнению обмена веществ. Первый период - развитие в яйцеводе и в первые 30-33 часа инкубации, питание и дыхание до появления кровеносной системы; второй - развитие с 33 часов до 5-го дня инкубации, питание и дыхание с момента образования желточного кровообращения до развития аллантоидного кровообращения; третий - с 6 по 19-й день инкубации, питание и дыхание при помощи желточной и аллантоидной систем кровообращения; четвертый - 20 и 21 дни инкубации, переход к дыханию при помощи легких.
Взяв за основу характер обмена веществ и тесно связанные с ним процессы кровообращения зародыша, авторы различают пять стадий:
I - стадия латебрального питания охватывает период от овуляции до организации желточного кровообращения (30-36 часов);
II - стадия желточного питания при помощи желточного круга кровообращения до 7-8-го дня инкубации;
III - стадия дыхания атмосферным кислородом и питания белком яиц от 7-8 до 18-19-го дня инкубации;
IV - стадия потребления зародышем кислорода воздушной камеры яйца с 18-19-го дня до наклева;
V - стадия 1-2-го дня от наклева до вывода.
Как утверждают авторы, в основу установления периодов и стадий положены одни и те же изменения обмена веществ. В обоих случаях не приняты во внимание условия, в которых осуществляется инкубация яиц.
Г. К. Отрыганьев в отдельные периоды эмбрионального развития видел изменения требований организма к условиям внешней среды. Это, по его мнению, дает возможность соответствующим режимом инкубации направленно влиять на зародыш. С начала дробления яйцеклетки в яйцеводе и до момента выхода цыпленка из яйца Г. К. Отрыганьев различает 11 периодов морфологических преобразований, которые соответствуют определенным физиологическим преобразованиям. Первый период проходит в яйцеводе птицы до снесения яйца. Второй период наступает с момента снесения яйца и характеризуется понижением до минимума всех физиологических процессов. Третий период, до 16-го часа инкубации,- активация роста и других физиологических процессов. Четвертый период, с 16 по 36-й час, - усложнение обмена веществ и появление новой плазмы. Пятый период, с 36 до 60-го часа,- начало кровообращения, дальнейшая дифференциация органов и начало отделения головы от бластодермы. Шестой период, с 21/2 до 51/2 дней - образование амниона, погружение зародыша в желток, интенсивный рост оболочек, накопление молочной кислоты. Седьмой период, с 51/2 до 10 дней,- охват аллантоисом внутренней поверхности скорлупы, переход на аллантоидное дыхание и испарение влаги частично за счет аллантоиса. Восьмой период, с 11 по 13-й день,- начало деятельности почек и пищеварительной системы. Девятый период, с 13 по 16-й день,- повышение интенсивности жирового обмена, увеличение внутрияйцевой температуры, появление способности к теплорегуляции. Десятый период, с 17 по 19-й день,- начало атрофии системы кровообращения аллантоиса, втягивание желтка. Одиннадцатый период, 20 и 21-й дни,- переход на дыхание легкими, выход из скорлупы.
Позже Г. К. Отрыганьев пришел к заключению, что следует разяичать как бы два периода, во время которых следует дифференцировать режим инкубирования: первый, когда яйцо требует интенсивногообогрева, и второй, когда в яйце образуется избыток физиологического тепла, который необходимо устранить.
На основании изучения роли белковой оболочки яйца в период развития цыпленка внутри скорлупы М. Н. Рагозина уточнила выдвинутые ранее Г. А. Шмидтом три периода; собственно зародышевый, предплодный и плодный. Зародышевый период начинается с момента оплодотворения яйца и заканчивается к концу седьмых суток развития зародыша при инкубации. За это время происходит закладка почти всех систем органов. Вес белковой оболочки значительно уменьшается вследствие перехода воды и растворимых в ней минеральных веществ в желток. Предплодный период продолжается с 8 по 12-й день инкубации. За это время происходит интенсивный органогенез, завершающийся формированием раннего плода. Вес белковой оболочки не убывает, так как жидкая часть ее была отдана в предыдущий период. Желточный мешок в предплодный период уменьшается в размерах и достигает своего первоначального веса.
В эти периоды зародыш использует для построения своего тела содержимое желточного мешка и жидкой части белковой оболочки. Усвоение веществ происходит при помощи специально образовавшейся системы ворсинок, энтодермального слоя желточного мешка, заменяющей собой систему ворсинок тонкой кишки взрослого организма.
Плодный период начинается на 13-й день инкубации и заканчивается в момент вывода цыпленка. Этот период связан главным образом с ростом организма. Белок начинает передвигаться через сероамниотический ход в полость амниона. Этому передвижению способствует секреция внутреннего клеточного слоя сероамниотического канала и белкового мешка (С. А. Шейнис). Белок из полости амниона заглатывается зародышем и усваивается внутрикишечно.
М. Н. Рагозина конкретизировала также периоды развития утки (зародышевый период 0-9 суток, предплодный 10-16 суток, плодный период 17-24 дня) и обнаружила короткий специфический период вылупления на 25-27-е сутки (у курицы па 20-21-е сутки).
Н. П. Бордзиловская периоды развития уток характеризует по существу в связи с различиями в способах питания эмбрионов. А. В. Денисьевский, изучая рост и развитие индейки, кладет в основу периодизации различия в строении органов, обеспечивающих поступление пищевых продуктов, и газообмен.
Г. П. Еремеев показал синхронность эмбрионального развития многих видов птиц, обусловливаемую общностью схемы строения яйца и его химического состава, малой изменчивостью относительного веса составных частей - плазм и скорлупы, сходством условий развития в яйце и приспособленностью к условиям внешней среды. Синхронность им устанавливалась по совпадению морфологических и физиологических изменений.
Явление синхронности зародышевого развития разных видов птиц имеет не только теоретическое значение, оно позволяет сделать важные практические выводы. Например: режим инкубирования всех видов сельскохозяйственных птиц в первую половину инкубации (примерно до замыкания краев аллантоиса) одинаков; видовые особенности, требующие изменения режима, приобретают значение главным образом во вторую половину инкубационного периода.
Для удовлетворения современных повышенных требований к результатам инкубации, как к средству размножения птицы и улучшения ее организма, для совершенствования технологии инкубирования, необходима дальнейшая углубленная разработка периодизации зародышевогоразвития птицы. Такая разработка должна быть направлена на совершенствование приемов управления развитием зародышей и контроля за его развитием. Важно установить зависимость периодичности развития зародыша как от качеств яйца, определяющихся в процессе его формирования и оплодотворения яйцеклетки, так и от условий, в которых находится яйцо во время инкубации. Под влиянием взаимосвязи этих двух моментов создается внутри яйца та среда, в которой осуществляется рост и развитие зародыша. Было бы неправильно изучать закономерности развития зародышей вне зависимости от условий, в которых находится яйцо во время инкубации.
Дыхание зародыша (часть вторая)
Исследования Л. А. Бражниковой дают представление о некоторых видовых особенностях обмена газов у яиц уток. В период с 9 на 26-й день инкубации яйцо утки выделяет 6602,7 см3 углекислоты и поглощает 9169,1 см3 кислорода, что соответствует дыхательному коэффициенту 0,727 в среднем за период. За последние 9 дней инкубации газообмен в утином яйце проходит на более высоком уровне, чем газообмен в курином яйце в последние 7 дней. В то же время за предыдущие 9 дней инкубации утиных и 7 дней инкубации куриных яиц газообмен в утиных яйцах ниже, чем в куриных.
Гусиное яйцо весом 165 г, как определил А. Ю. Быховец, за 30 дней инкубации выделяет 14 017,2 см3 кислорода, что в среднем соответствует дыхательному коэффициенту 0,728.
Л. М. Баранчеев, исследуя дыхание эмбрионов яиц кур и уток во время инкубации, установил закономерное снижение дыхательного обмена, совпадающее с депрессией роста зародыша, интенсивностью морфогене-тических процессов и сменой типа обмена веществ. Л. А. Бражникова также нашла, что снижение скорости роста зародыша утки связано со снижением интенсивности обмена веществ.
Л. М. Баранчеев, А. Ю. Быховец и Л. А. Бражникова показали, что интенсивность газообмена - выделение углекислоты на единицу веса зародыша яиц кур, уток и гусей уменьшается к концу инкубации, что соответствует снижению скорости роста зародыша. Л. М. Баранчеев наблюдал уменьшение поглощения кислорода яйцами, в которых зародыши погибают при выводе: у яиц кур понижение дыхательного обмена наблюдается с 13-го, а у яиц уток - с 17-го дня инкубации.
Дыхательный коэффициент в инкубируемых яйцах у птицы всех видов близок к 0,7. Общим является и то, что в первые дни инкубации дыхательный коэффициент приближается к единице. Это указывает на углеводный характер обмена веществ. Затем дыхательный коэффициент снижается до уровня, характерного сначала для протеинового, а затем и жирового обмена.
Нидхэм показал, что дыхательный коэффициент бластодермы в первые два дня характерен для углеводного обмена, но для area opaca, взятой отдельно, дыхательный коэффициент падает до 0,7-0,8. С 3 по 6-й день дыхательный коэффициент зародыша неизменно равен 1,0. После этого он понижается и достигает к концу инкубации уровня, характерного для протеинового, а затем жирового обмена (около 0,7). Дыхательный коэффициент только что выведенного цыпленка быстро повышается и К 5-му дню его жизни приближается к единице. Дыхательный коэффициент желточного мешка быстро снижается и в последние дни 'инкубации бывает ниже 0,6. Дыхательный коэффициент аллантоиса снижается медленно и сохраняется на уровне около 0,8. Нидхэм на основе исследований Тангля, Митуха, Бора, Хас-сельбаха и Мюррея сделал расчет энергетических процессов во время инкубации куриного яйца. Согласно этим расчетам, энергетические процессы в яйце во время инкубации можно представить в следующем виде:
|
- |
Калорий |
% |
|
В яйце содержиться до инкубации |
88,9-89,0 |
- |
|
Остаеться в желточном мешке |
41,0-27,0 |
45,4-31,0 |
|
Содержиться в зародыше |
32,0-37,0 |
36,4-42,5 |
|
Потеря за период инкубации (вследствие окисления) |
16,0-23,0 |
18,2-26,4 |
Общая калорийность зародыша увеличивается по мере его роста (Э. Э..Пенионжкевич и Л.И.Шехтман). Вместе с тем увеличивается и калорийность 1 г сухих веществ в связи с относительным уменьшением неорганической части и повышением содержания жира. Так, в 1 г сухого вещества белка и желтка содержится 6,94 калории, в 1 г эмбриона на 5-й день инкубации - 5,1 калории и в 1 г сформировавшегося цыпленка - 6,2 калории.
Тепло, освобождающееся в яйце вследствие окисления питательных веществ, оказывает влияние на температуру внутри яйца. Э. Э. Пенионжкевич и А. Н. Ретанов установили два термических периода. Первый период начинается с момента помещении яйца в инкубатор, а второй - через 9 суток и 3 часа после закладки яйца в инкубатор и заканчивается после вывода цыпленка. В первый период температура внутри яйца, как правило, ниже температуры воздуха инкубатора или равна ей, редко бывает выше нее.
Второй термический период внутри яйца характеризуется тем, что изменяющаяся температура в яйце не опускается ниже температуры воздуха инкубатора и имеет тенденцию к повышению к концу инкубации. Температура внутри яйца достигает максимума в последний день инкубации, она равна 45,9°, или на 6,8° выше температуры в инкубаторе. Аналогичные результаты получила С. Л. Слинько.
Ромейн также нашел, что температура яйца после 10 дней инкубации равна или несколько ниже температуры окружающего воздуха. Температура яйца через 13 дней инкубации уже заметно выше внешней температуры, и на 19 и 20-й день инкубации эта разница равнялась 1-2°.Из сопоставления этих работ можно сделать вывод, что не только режим инкубатора определяет уровень температуры внутри яйца, последняя зависит и от интенсивности обмена веществ.
В опыте Э. Э. Пенионжкевича и А. И. Ретанова развитие зародыша проходило более интенсивно, и уже в первый период температура яйца была более высокой и близкой к температуре воздуха инкубатора. Во второй период температура яйца была много выше температуры воздуха инкубатора. Можно предположить, что в связи с более интенсивным развитием зародыша температура внутри яйца в этом опыте значительно раньше превысила температуру воздуха инкубатора, чем в других опытах. Еще более раннее повышение температуры яиц наблюдал Романов при температуре в инкубаторе 37,5°.
Прижизненная оценка развития зародышей (часть вторая)
При плохом развитии зародыш мал, не погружен в желток, расположен близко к скорлупе и хорошо виден, особенно его глаз. Кровеносная система мало развита, и желточные кровеносные сосуды подходят к самому зародышу. Вокруг зародыша светлого поля не заметно (рис.1, А).
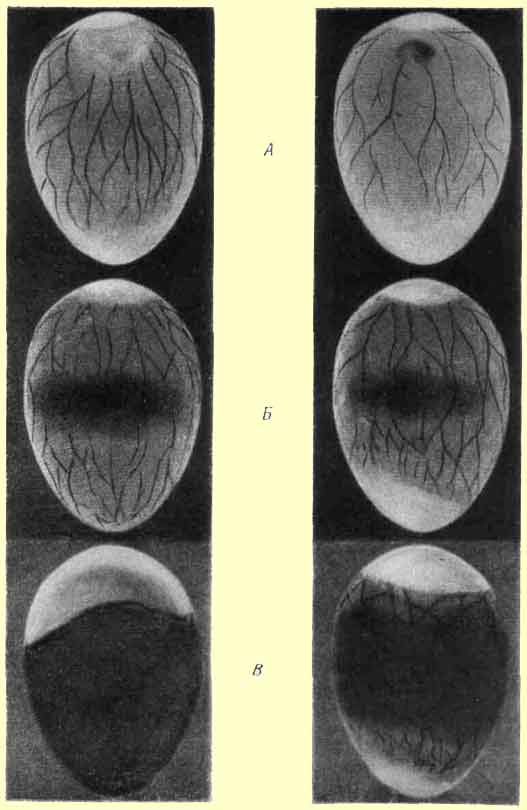
Рис.1.Просвеченные яйца. Справа - яйца с хорошо развитыми зародышами,слева - яйца с отстающими в развитии зародышами. А - через 6 дней инкубации, Б - через 11 дней, В - через 19.
Разница в результатах инкубации яиц с различным характером развития зародышей на 6-й день инкубации показана в таблице 11 (по данным М. В. Орлова и Е. Н. Кучкрвской).
Таблица 11. Вывод цыплят из яиц с разным характером развития зародышей на 6-й день инкубации (в %)
|
№ опыта |
Яйца с хорошо развитыми зародышами |
Яйца с отставшими в развитии зародышами |
Разница в проценте вывода |
|
I |
51,2 |
32,4 |
18,8 |
|
II |
78,3 |
54,6 |
23,7 |
|
III |
92,9 |
89,6 |
3,3 |
После 11 суток инкубации при просвечивании яиц с хорошо развитыми зародышами аллантоис выстилает всю скорлупу внутри яйца, охватывает весь белок и замыкается в остром конце яйца. Иногда между его краями имеется очень небольшое расстояние, которое вскоре закрывается. Зародыш крупный, темный.
Если зародыш отстает в развитии, то задерживается и рост аллантоиса; около острого конца яйца видна граница окрашенного в розовый цвет аллантоиса, а в остром конце яйца просвечивается светлый белок; зародыш мал (рис.1, Б).
В таблице 12, но данным М. В. Орлова и Е. Н. Кучковской, приведены результаты вывода цыплят из яиц с замкнутым и незамкнутым аллантоисом на 11-и день инкубации.
Таблица 12. Вывод цыплят из яиц с замкнутым аллантоисом (в %)
|
№ опыта |
Яйца с замкнутым аллантоисом |
Яйца с незамкнутым аллантоисом |
Разница в проценте вывода |
|
I |
64,7 |
26,4 |
38,3 |
|
II |
84,5 |
46,4 |
38,1 |
|
III |
96,0 |
84,8 |
11,2 |
После 19 суток инкубации при третьем просмотре крупный зародыш заполняет все яйцо и острый конец его не просвечивается. Зародыш выпячивает в воздушную камеру свою шею, видна подвижная тень ее. Воздушная камера обычно имеет достаточно большие размеры, но иногда может быть и невелика.
При отсталом развитии яйцо просвечивается как в остром конце, так и около воздушной камеры; зародыш мал и не выпячивает своей шеи в воздушную камеру. Воздушная камера обычно небольшая (рис.1, В).
Режим инкубирования (часть вторая)
В таблице 6 приведены данные, характеризующие развитие зародышей в яйцах, одновременно заложенных в инкубатор "Рекорд-39"различным образом (М. В. Орлов). Яйца контрольных групп закладывали по схеме инкубатора "Рекорд-39": сверху и снизу закладываемой партии были яйца с зародышами, имевшими к этому времени возрасг 9-12 дней. Яйца опытных групп закладывали по схемам инкубаторов "Коммунар" и "Укргигант": три или два ряда лотков закладывали сверху подряд и под последним рядом были яйца, заложенные на три дня раньше.
Таблица 6. Результаты инкубации яиц, размещенных по схемам разных инкубаторов
|
I опыт |
I опыт |
II опыт |
II опыт |
|
|
Инкубатор "Рекорд 39" |
Инкубатор "Коммунар" |
Инкубатор "Рекорд 39" |
Инкубатор "Укргигант" |
|
|
Показатели |
контрольные |
подопытные |
контрольные |
подопытные |
|
Рост и развитие зародышей в первые два дня: |
||||
|
- диаметр сосудистого поля, мм |
11,4 |
9,2 |
10,7 |
10,2 |
|
- длина зародыша, мм |
5,8 |
5,7 |
5,6 |
5,4 |
|
- количество пар сомитов |
18,5 |
16,1 |
17,2 |
16,8 |
|
Средняя продолжытельность инкубации (в часах) |
488,1 |
493,3 |
488,4 |
492,1 |
|
Вывод здоровых цыплят, % |
93,5 |
90,7 |
93,6 |
89,5 |
Как видно из таблицы 6, развитие и рост зародышей в яйцах, заложенных по схеме инкубатора "Рекорд-39", были лучше, чем в яйцах подопытных партий. После двух дней инкубации яйца подопытных партий были размещены по схеме инкубатора "Рекорд-39" и находились в таком положении до конца инкубации. Хорошее развитие зародышей в яйцах контрольных партий в первые дни инкубации оказало благоприятное влияние на весь последующий ход инкубации и вывод молодняка был в них более своевременный и более успешный.
Ромейн и Локхорст (Romijn G. and W. Lokhorst) в эксперименте в "темной камере" с искусственным зародышем в яйце определили, что нл. 18-й день инкубации яйцо курицы теряет 150 кал/час. Из этого количества на испарение воды расходуется 11 кал/час, проведением теряется 65,7 кал/час, а путем радиации 73,3 кал/час, или почти половина всего количества тепла.
Из сказанного следует, что в шкафных инкубаторах, когда рядом с закладываемыми яйцами находятся пустые лотки, для обеспечения успешного развития зародышей необходимо вначале поддерживать температуру более высокую, чем обычно: 37,7-37,8° вместо 37,4-37,5°.
Закладка яиц по схеме инкубатора "Рекорд-39" создает благоприятные условия отдачи излишнего тепла яйцами и во вторую половину инкубации.
Существенно изменяет влияние температуры воздуха инкубатора влажность воздуха. Г. К. Отрыганьев и Е. Н. Кучковская показали, что низкая влажность в секционном инкубаторе при повышенной температуре вызывает резкое повышение смертности зародышей в первые 5 дней инкубации. Высокая же влажность в этих условиях значительно снижает смертность зародышей. Авторы сделали вывод, что в первые дни инкубации наиболее благоприятное влияние оказывает сочетание высокой температуры с высокой влажностью, а наименее благоприятно-сочетание низкой температуры с высокой влажностью.
Н. А. Коноплев отмечает, что рост и развитие зародыша в шкафных инкубаторах протекают интенсивнее при более высокой влажности* По его данным, повышение температуры с 36,5 до 38,5° при влажности 40% задерживает развитие зародыша, а при влажности 80% обычно ускоряет развитие.
Анализ динамики смертности зародышей (часть вторая)
Увеличение продолжительности хранения яиц приводит к резкому увеличению смертности зародышей в первые дни развития, как это видно из данных таблицы 18 (М. В. Орлов).
Таблица 18. Распределение смертности зародышей курицы в зависимости от продолжительности хранения яиц перед инкубацией (в % от общего количества мертвых зародышей за весь период инкубации)
|
Количество мертвых зародышей |
Продолжительность хранения яиц (в днях) |
Смертность с 1 по 6 день |
Смертность с 7по 15 день |
Смертность с 16 по 21 день |
|
98 |
1 |
31,0 |
20,0 |
49,0 |
|
309 |
15 |
84,4 |
5,0 |
10,6 |
|
424 |
25 |
99,7 |
0,0 |
0,3 |
Э. Э. Пенионжкевич наблюдал, что увеличение срока хранения яиц вызывает гибель зародыша еще до закладки в инкубатор и на первых стадиях эмбрионального развития, это приводит к увеличению "условно неоплодотворенных" яиц. То же самое наблюдал и И. Я. Прицкер. Поэтому можно считать, что резкое увеличение смертности зародышей в первые дни инкубации и увеличение количества "неоплодотворенных" яиц, которые при вскрытии оказываются оплодотворенными - специфично для старых и плохо хранившихся яиц.
С неполноценностью яиц, вызванной недостаточностью в рационе несушек витаминов, главным образом группы В, или неполноценностью протеинов, связано повышение смертности зародышей в средние дни инкубации. Нарушение водно-минерального и протеинового обмена может привести к тому, что количество мертвых зародышей может быть не только больше количества кровяных колец, но и больше количества задохликов.
В таблице 19 приведены результаты инкубации яиц кур, в рационе которых в январе и феврале было недостаточно рибофлавина и биотина. Эти же куры в июне и июле получали полноценные корма и пользовались выгулом.
Таблица 19. Распределение мертвых зародышей в зависимости от степени полноценности инкубационных яиц (в % от всех умерших зародышей)
|
Месяц |
Количество проинкубированных яиц, шт |
Вывод, % |
Смертность с 1 по 6 день |
Смертность с 7 по 18 день |
Смертность с 19 по 21 день |
|
Январь - Февраль |
209390 |
48,3 |
20,1 |
56,8 |
23,1 |
|
Июнь - Июль |
42392 |
80,1 |
40,0 |
31,4 |
28,4 |
На основании этих данных можно сделать вывод, что значительное увеличение смертности зародышей в средние дни инкубации специфичнодля неполноценных яиц (следствие неполноценности рациона маточного стада).
Резкие нарушения протеиновой части рациона вызывают также увеличение количества мертвых зародышей. Следует отметить, что чем острее недостаток витаминов в яйце, тем раньше наступает гибель зародышей.
В практической работе необходимо в каждом случае установить, чем вызвана неполноценность яиц (неправильное кормление маточного стада, неудовлетворительное хранение яиц) или какие и когда были допущены ошибки в режиме инкубирования.
В дальнейшей диагностической работе основное значение приобретают признаки, указывающие на характер расстройств развития зародышей, на патологию развития. Однако встречаются серьезные трудности, связанные с тем, что патологические процессы в яйце возникают постепенно, проходят определенные этапы, нарушая нормальный ход развития, и приводят организм к смерти. Здесь не один процесс реакции организма на патогенный фактор, но и быстрый процесс онтогенеза, в различные моменты которого реакции организма различны и связаны с нарушением дальнейшего развития.
Вполне возможно, что не только в разном возрасте эмбрион по-иному реагирует на внешний фактор, но и различные факторы могут вызвать у него одинаковую реакцию. 6 связи с этим далеко не все патологические отклонения являются специфичными для фактора, вызвавшего эти отклонения. Кроме того, в каждой партии инкубируют яйца, существенно отличающиеся по своим свойствам и поэтому по-разному реагирующие на одни и те же условия инкубации.
И. Я. Прицкер отмечает, что при отклонениях температуры инкубации на несколько десятых градуса от нормы вполне отчетливой и ясно выраженной специфичности поражений эмбрионов не наблюдается. Поэтому для установления причин неудовлетворительных результатов инкубации важны не отдельные поражения, а частота их повторений, встречающаяся среди вскрываемых яиц с мертвыми зародышами.
Для того чтобы иметь представление о патологии мертвых зародышей, характерной для данной партии яиц, необходимо вскрыть от 5 до 10% яиц из каждой категории отходов.
Для диагностики имеет значение классификация эмбриональных заболеваний, разработанная Г. К. Отрыганьевым.
Патологические процессы:
- Экзогенные
----- Старение яйца (заражение яйца - аспергиллез)
----- Высокая температура (аморфоз, бластодермальный кистоз, уродства головы, эктопия, кистоз эмбриональной оболочки, гипертермическая кахексия, острая гипертермия)
----- Низкая температура (гипотермический энтерит, гиперкардия)
----- Избыточная влажность (амниогенная липкость, гидрогенная асфиксия)
----- Недостаточная влажность (аллантоисная геморрагия, агидроз)
----- Механические факторы (агидроз, агемезия аллантоиса)
----- Недостаточная вентиляция (экзогенная асфиксия)
- Эндогенные
----- Патология гамет
----- Патология оплодотворения
----- Патология яйцевых плазм и оболочек (абиотиноз, арибофлавиноз, акобаламиноз, микседематоз, эмбрион. подагра, авитаминоз А, авитаминоз Е, кутикулит)
----- Заражение яйца (ББП, паратиф, омфолит, вирусный гепатит, аспергиллез).
Диагностика неполноценности яиц (часть вторая)
В тканях зародыша откладывается большое количество мочекислых солей и фосфорнокислого кальция. Чаще всего поражаются почки: они увеличены, пронизаны белыми кристаллами, достигающими величины макового зерна. При сильной степени заболевания кристаллы откладываются на перикарде, брыжейке и даже на желточном мешке (рис. 4, б).
Авитаминоз В2. При недостатке в рационе кур рибофлавина они несут яйца с бесцветным разжиженным белком, вследствие чего желток сильно подвижен и часто опущен к острому концу яйца.

Рис. 4. а - недоразвитое оперение у куриного зародыша, б - кальцинация почек зародыша, в - перозис, скользящий сустав, г - подкожная отечность, д - "попугаев клюв"
Развитие зародышей в таких яйцах, особенно во второй половине инкубации, задерживается. У части яиц аллантоис не замыкается в остром конце яйца или замыкается с опозданием. Обмен веществ нарушается, яйца мало испаряют воду, белок используется плохо, даже если он проникает в полость амниона. Вследствие этого выводятся "липкие" цыплята (Ю. М. Огородний и Э. Э. Пенионжкевич).
При просвечивании яиц перед выводом, помимо большого количества мертвых зародышей, обращает внимание сильное отставание их в росте, просвечивается большой участок в остром конце яйца, причем белок часто не покрыт аллантоисом.
Мертвые зародыши имеют характерный вид: шея их искривлена, конечности часто укорочены и пальцы искривлены, кожа покрыта нераскрывшимися перовыми сосочками ("булавовидный пух"), под подбородком наблюдается отек (рис. 4, а).
Поражение, связанное с авитаминозом В2, Г. К. Отрыганьев называет гиперкератозной формой дистрофии.
Недостаточность биотина.
При недостаточности биотина в рационе маточного стада признаки неполноценности яиц такие же, как и при авитаминозе В2. Отсутствие в рационе птицы биотина вызывает те же поражения, что и нарушение белкового питания (микромелическая форма дистрофии) (Г. К. Отрыганьев).
Отсутствие или недостаточность биотина начинает проявляться после начала использования белка зародышем непосредственно через рот. Это вызывает сильное повышение смертности зародышей, в особенности на 15-16-й день инкубации и при выводе.У погибших зародышей укороченные конечности, особенно задние, суставы утолщены, трубчатые кости искривлены. Резкой деформации подвергается голова: череп становится широким, укороченным, верхняя часть развивается больше нижней и загибается над ней вниз, придавая клюву "попугаеву форму" (рис. 4, д).
Мертвые зародыши покрыты липкой жидкостью амниона. Корпус зародыша мал, но зародыш кажется крупным: имеет большую голову и шарообразную форму вследствие втягивания большого, мало использованного желтка.
Вывод запаздывает и продолжается долго. Выведенные цыплята слабы, имеют большие животы. Как и при авитаминозе В2, смертность выведенных цыплят повышена (Коуч и др.).
У выведенных цыплят иногда наблюдаются атаксия и перозис (рис. 4, в). Атаксия выражается во втягивании головы у одних и подво-рачивании головы у других цыплят. Иногда у цыплят имеют место непрерывные вращательные движения головы, продолжающиеся до тех пор, пока не наступит смерть.
Пальцы у цыплят иногда повернуты назад, коленные суставы жесткие, и цыплята не могут стоять.
Недостаток витамина В12.
В опыте с курами, получавшими синтетический рацион, Олкез и др. (Olcese et al.) наблюдали при недостатке витамина В12 повышение смертности в период от 16 до 18-го дня инкубации. Наиболее характерным поражением была атрофия мышц ног, названная автором миоатрофия. Ноги были тонкие, почти нормальной длины и геморрагичные. Кроме того, наблюдались частые кровоизлияния зародыша и аллантоиса, а также необычно часто встречалось положение "голова между бедрами".
Недостаток фолиевой кислоты.
Смертность зародышей при недостатке фолиевой кислоты особенно велика в последние дни инкубации и даже после проклева скорлупы (Сунде и др. - Sunde et al.).
При остром недостатке фолиевой кислоты наблюдаются карлико-вость, ненормальности ног и клюва. Некоторые зародыши имеют искривленную берцовую кость (плюсна деформируется редко). Голова сплющенная, глаза маленькие и часто имеют прозрачный мешок над хрусталиком. Нижняя челюсть неразвита или отсутствует. Наблюдаются случаи общей отечности различной степени. Шея длинная и обычно скрученная. Органы брюшной полости растут быстрее, чем остальные.
Недостаток марганца.
При недостатке в рационе несушек марганца нарушается пористость скорлупы яйца и уменьшается ее прочность. Выводимость уменьшается при повышении смертности зародышей на 20-21-й день инкубации.
При недостатке марганца появляются симптомы микромиелии, выводятся цыплята с короткими ногами и крыльями, большой головой, попугаевым клювом, прямой (tibia) берцовой костью.
Часты случаи атаксии, выведенные цыплята вращают головой, запрокидывают ее на спину или подворачивают под туловище.
Следует заметить, что причиной появления атаксии может служить и недостаток тиамина, витамина В1 (аневрина). Подобные признаки наблюдаются и при недостатке биотина.
Ярким признаком недостаточности марганца является перозис, или, как его называли, скользящий сустав. Наиболее характерными симптомами этой болезни недостаточности, при которой нарушается развитие костей, являются: увеличенный предплюсневый сустав, искривление нижнего конца болыпеберцовой кости и верхнего конца плюсны, а также смещение ахиллесова сухожилия. Передвижение цыплят сильнозатруднено. При сильном поражении они не могут подобрать под себя ноги и не в состоянии встать. В таком случае они передвигаются порывисто на суставах, помогая себе крыльями.
Недостаток других минералов.
Значительное количество селена в кормах, выращенных в некоторых районах, вызывает снижение выводимости: смертность зародышей увеличивается, появляются уродства среди мертвых зародышей. Для отравления селеном характерна отечность и "проволочный" пух (жесткий).
Значение некоторых микроэлементов для развития зародышей было выяснено путем введения их в яйцо, главным образом в желток, в первые 4-8 дней инкубации.
Инъекция талия привела к появлению в 90-100 случаях ахондро-плязии. Слабая ахондроплязия появлялась и после введения в яйцо хрома. Олово и кобальт вызывают нарушения мозга. Бледность, отечность, а также задержка развития пуха были -следствием влияния бора. Мышьяк был причиной задержки роста зародышей, появления микро-мелии и отечности в брюшной полости. Примерно такие же нарушения наблюдались и после инъекции родия. Барий нарушал развитие ног (Карнофский).
Диагностика ошибок режима инкубирования (часть вторая)
По данным Г. К. Отрыганьева и Е. Н. Кучковской, перегрев в первые дни инкубации вызывает уродства головы: акранию, отсутствие или недоразвитие глаз, уродства лицевых костей и др. (рис. 2, а).
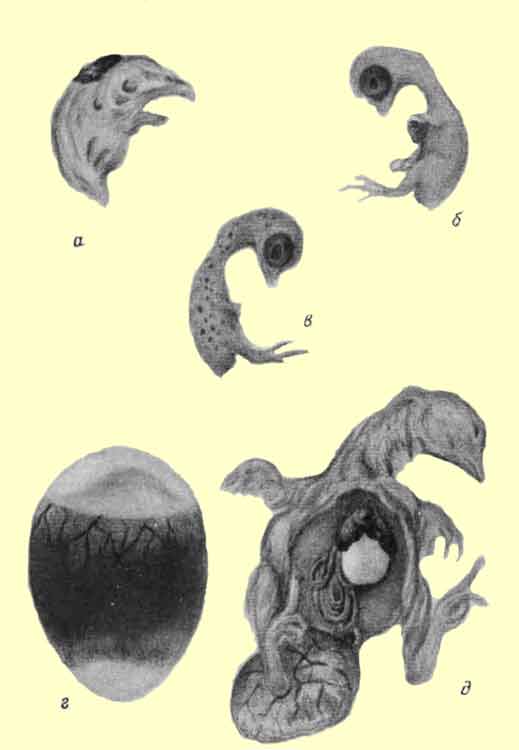
Рис. 2. Признаки перегрева. а - уродства головы, б - эктопия, в - подкожные кровоизлияния, г - куриное яйцо, просвеченное на 20-й день инкубации после длительного перегрева, д - вскрытый задохлик при перегреве.
С 3 по 5-й день инкубации перегрев вызывает нарушение отделения зародыша от желтка и появление эктопии (рис. 2, б).
Для перегрева зародыша в первые дни инкубации характерны гиперемии и кровоизлияния как в различных частях тела, так и в сосудистом поле на желтке. Очень часто встречаются прилипания зародыша и бластодермы к скорлупе.
Во время просвечивания яиц после 6 суток инкубации при перегреве наблюдается большая неоднородность зародышей: наряду с очень хорошо развитыми, глубоко лежащими, имеется значительная часть неудовлетворительно и плохо развитых зародышей и большое количество ярко выраженных кровяных колец.Перегрев в средние дни инкубации не вызывает специфических нарушений в развитии уже сформировавшегося зародыша и его оболочек. Но Г. К. Отрыганьев отмечает, что при перегреве в период с 9,5 по 12,5 день стенки амниона и аллантоиса часто покрыты прозрачными пузырьками - кистами различной величины. Очень сильное повышение температуры в средние дни инкубации приводит к смерти большого количества зародышей. При этом кровеносные сосуды аллантоиса переполняются кровью, в коже зародыша, иногда в мозге и сердце, появляются точечные кровоизлияния (рис. 2, б), характерна также гиперемия печени, мозга и почек.
Признаки перегрева очень характерны в последние дни инкубации и при выводе. Высокая температура задерживает использование белка и желтка, и в связи с этим зародыши имеют относительно малый вес (М. В. Орлов). При просвечивании яиц на 19-й день инкубации обнаруживается, что многие зародыши уже выпячивают шею в воздушную камеру, но в то же время в остром конце просвечивается остаток покрытого аллантоисом белка (рис. 2, г). Наряду с этим имеются яйца с некоторыми признаками отставания в развитии зародыша.
Наклев начинается рано, скорлупа отламывается маленькими кусочками. Много яиц с "сухим" наклевом в остром конце яйца. Выведенный молодняк мелкий, плохо опушен, имеет грубый большой живот. Пуповина часто плохо заживлена, на ее месте остается подсохшая кровь, а у части молодняка кровоточит пуповина.
Скорлупа, оставшаяся после вывода, имеет темно-красный или бурый цвет из-за переполнения кровью сосудов аллантоиса. Часто можно наблюдать кровоизлияние в аллантоисе. В скорлупе на остром конце яиц часто остается неиспользованный белок.
Все невыведшиеся зародыши мертвы и многие из них имеют неправильные положения и кривые пальцы на ногах. В яйцах наблюдаются остатки плотного неиспользованного белка, зародыши не втянули большой гиперемированный желточный мешок.
У вскрытых задохликов обнаруживается гиперемия полостей, кишечника, сердца (Г. К. Отрыганьев и Г. И. Крылов, Г. К. Отрыганьев). Сердце часто уменьшенных размеров (И. Я. Прицкер и Е. Ф Лисиц-кии) (рис. 2, д).
Влажность (часть вторая)
Сразу же после снесения яйцо начинает терять вес. Потеря веса особенно увеличивается после начала инкубации. Считают, что вес яйца уменьшается только вследствие испарения воды. На самом деле истинный вес воды, испарившейся из яйца, несколько больше потери веса яйцом, так как вес поглощенного кислорода больше веса выделенной углекислоты. Только что снесенное яйцо не содержит излишков воды; поэтому при потере ее до начала инкубации ухудшаются условия жизни .зародыша (М. В. Орлов).В таблице 5 приведены некоторые данные о потере веса яйцами. За период инкубации яйца кур теряют 6-7 г, яйца индеек - 9-11 и яйца гусей - 16-28 г.
Таблица 5. Потеря веса яйцами во время инкубации
|
Вид птицы |
За сколько дней |
Потеря веса, % |
Вывод молодняка, % |
Способ вывода |
Чьи данные |
|
Куры |
20 |
11,1 |
100,0 |
Под наседкой |
В.В.Фердинандов |
|
-- |
19 |
15,8 |
89,0 |
-- |
Н.П.Третьяков |
|
-- |
19 |
13,8 |
67,2 |
Секционный инкубатор |
-- |
|
-- |
18 |
13,5 |
100,0 |
Под наседкой |
Э.Э.Пенионжкевич, Л.М.Шехтман |
|
-- |
19 |
12,9 |
89,7 |
Шкафный инкубатор |
А.Э.Пенионжкевич |
|
-- |
19 |
12,8 |
90,0 |
-- |
-- |
|
-- |
19 |
12,1 |
90,2 |
-- |
-- |
|
Индейки |
28 |
12,8 |
77,8 |
-- |
А.У.Быховец |
|
-- |
25 |
13,2 |
81,9 |
-- |
-- |
|
-- |
24 |
15,4 |
66,1 |
Секционный инкубатор |
М.В.Орлов |
|
-- |
24 |
15,1 |
71,8 |
-- |
-- |
|
-- |
24 |
15,2 |
79,0 |
-- |
-- |
|
-- |
25 |
11,7 |
69,9 |
Шкафный инкубатор |
Н.П.Третьяков |
|
-- |
25 |
14,0 |
85,4 |
Под наседкой |
-- |
|
Утки |
24 |
8,6 |
70,3 |
Шкафный инкубатор |
А.Э.Пенионжкевич |
|
-- |
24 |
13,1 |
89,7 |
-- |
-- |
|
-- |
24 |
15,9 |
95,6 |
-- |
-- |
|
Гуси |
28 |
15,7 |
69,7 |
Секционный инкубатор |
Л.А.Бражникова |
|
-- |
27 |
16,4 |
71,7 |
-- |
-- |
|
-- |
27 |
13,2 |
78,3 |
-- |
-- |
|
-- |
30 |
12,8 |
80,6 |
Под наседкой |
-- |
|
-- |
30 |
13,1 |
66,9 |
Шкафный инкубатор |
-- |
|
-- |
27 |
12,5 |
59,0 |
-- |
Ю.Н.Владимирова |
|
-- |
27 |
10,2 |
62,3 |
-- |
-- |
|
-- |
27 |
10,9 |
65,4 |
-- |
-- |
Испарение воды оказывает большое влияние на общий баланс тепла в яйце. За время инкубации на испарение воды расходуется 15-20% тепла, образующегося в яйце. Потеря тепла в связи с испарением воды имеет особенно большое значение в начале инкубации, когда эта потеря очень мало может быть компенсирована за счет незначительного количества окисляемых веществ.
Влажность воздуха в инкубаторе оказывает влияние на отдачу тепла: яйцо теряет больше тепла во влажном воздухе, так как теплопроводность воздуха влажного выше теплопроводности сухого. Это имеет большое значение во вторую половину инкубации, когда в яйце при хорошем развитии зародыша температура выше, чем температура воздуха инкубатора.С точки зрения обогревания яйца высокая влажность в инкубаторе оказывает благоприятное влияние как в начале, так и в конце инкубации. В первые дни инкубации это связано с уменьшением испарения воды и сохранением в связи с этим тепла в яйцах, что способствует хорошему развитию зародышей и уменьшает их смертность.
Пониженная влажность в отот период вызывает усиленное испарение воды, что ухудшает условия обогревания яиц. При уменьшении запасов воды в яйцах затрудняется растворение питательных веществ и поступление их к зародышам. Вследствие этого замедляется их рост и возрастает смертность.
В последние дни инкубации высокая влажность усиливает теплоотдачу, предупреждая перегрев яиц и вызывая такую же положительную реакцию хорошо развитых зародышей, как и при некотором снижении температуры.
Потеря в весе при одинаковой относительной влажности разных яиц может значительно отличаться. По данным Ю. Н. Владимировой, яйца молодок теряют в весе больше, чем яйца переярых кур. Яйца, снесенные летом, теряют в весе больше, чем снесенные зимой. Мелкие яйца также теряют относительно больше, чем крупные.
Чем больше теряют яйца в весе до инкубации, тем интенсивнее испаряется вода из них во время инкубации. После длительного хранения яйца очень много теряют в весе в первые дни инкубации и мало испаряется из них воды во вторую половину инкубации (М. В. Орлов, 1948). Было установлено несколько периодов в жизни зародыша, когда он различно реагирует на условия влажности воздуха.
- С 1 по 6-й день инкубации, когда белок яйца не защищен аллантоисом, испарение воды в яйце зависит главным образом от влажности воздуха инкубатора. Потеря воды, еще не использованной зародышем, в это время невозвратима. Регулирующая роль влажности в отношении потери тепла яйцом в этот период осуществляется в связи с испарением воды.
- С 6 по 10-11-й день, когда аллантоис постепенно покрывает весь белок и замыкается в противоположном от зародыша конце яйца, испарение воды белка постепенно заменяется потерей воды аллантоиса, которая уже принимала участие в обмене веществ.
- С 10-11 по 15-й день вода испаряется исключительно из аллантоиса, полностью закрывающего все содержимое яйца. Влажность воздуха в этот период уже не оказывает такого же влияния на испарение-воды из яиц, как в первые дни. Все большее значение в этом процессе приобретает интенсивность обмена веществ в яйце.
- С 16-го дня и до конца инкубации, когда постепенно отмирает аллантоис, испарение воды очень мало зависит от относительной влажности воздуха инкубатора. Регулирующая роль влажности в отношении потери тепла яйцом обусловливается изменением теплоемкости и теплопроводности воздуха инкубатора, имеющего более низкую температуру.
Инкубация в современных шкафных инкубаторах производится при относительной влажности воздуха в пределах от 45% до 65-70%. и выше.
Режим инкубирования (часть третья)
В той же работе Н. А. Коноплев получил материал, характеризующий значение скорости движения воздуха в комплексе внешних условий (скорость движения воздуха измерялась между колонками лотков). Результаты его опыта первых 2 дней инкубации приведены в таблице 7.
Таблица 7. Влияние различных сочетаний температуры, относительной влажности и скорости движения воздуха (в м/с) на количество сомитов куриного зародыша в первые 48 часов инкубации
|
Влажн. 40% |
Влажн. 40% |
Влажн. 40% |
Влажн. 40% |
Влажн. 80% |
Влажн. 80% |
Влажн. 80% |
Влажн. 80% |
|
|
Температура,° |
Скорость 0,5 м/с |
Скорость 0,7 м/с |
Скорость 1,6 м/с |
Скорость 2,0 м/с |
Скорость 0,5 м/с |
Скорость 0,7 м/с |
Скорость 1,6 м/с |
Скорость 2,0 м/с |
|
36,5 |
1,7 |
15,9 |
15,2 |
15,9 |
-- |
15,5 |
20,6 |
-- |
|
37,5 |
15,2 |
14,5 |
13,9 |
19,9 |
18,6 |
18,7 |
19,8 |
-- |
|
38,5 |
12,2 |
13,8 |
14,8 |
18,1 |
23,7 |
29,2 |
29,6 |
28,3 |
Как видно из таблицы, пониженная температура воздуха задерживает развитие зародыша тем сильнее, чем больше скорость движения воздуха (от 0,5 до 1,6 м/сек). Повышение температуры при высокой влажности ускоряет развитие зародыша тем сильнее, чем больше скорость движения воздуха. Увеличение скорости движения воздуха при низкой температуре и влажности задерживает, а при высокой температуре ускоряет развитие; увеличение скорости движения воздуха при высокой влажности ускоряет развитие как при низкой, так и при высокой температуре. Точно также изменяются размеры сосудистого поля и зародыша.
Температура (часть вторая)
М. В. Орлов на основании своих работ пришел к выводу, что небольшое повышение температуры в первые дни инкубации обеспечивает ускорение роста зародыша и оказывает благоприятное влияние на весь период его развития и конечный результат инкубации; понижение температуры после 15-го дня инкубации создает лучшие условия для окончания роста зародыша. Это положение было подтверждено работой Г. С. Кот-лярова, который, проинкубировав 6,5 тыс. яиц в секционном инкубаторе, пришел к выводу, что целесообразно небольшим повышением температуры в начале инкубации вызвать ускорение развития зародышей, а путем понижения температуры (увеличение теплоотдачи) улучшить условия для вывода.
Температура воздуха инкубатора оказывает существенное влияние на интенсивность развития зародыша. Пониженная температура задер-живает развитие зародыша и может увеличить продолжительность инкубации в среднем на четверо суток. Повышенная температура уменьшает продолжительность инкубационного периода в среднем менее чем на одни сутки. Очень высокая температура задерживает вывод молодняка (Романов и др.). Наибольшее влияние на продолжительность инкубации оказывает температура в первые дни инкубации, меньшее - в средние дни и почти не оказывает влияния в последние дни, если инкубируют яйца с хорошо развитыми зародышами.
В шкафном инкубаторе типа КЭМ-20 М. Ф. Сорока изменил продолжительность инкубационного периода яиц уток, поддерживая во вторую половипу инкубации температуру от 36,5 до 38,5°. По его данным, повышение температуры на 1° сокращало в среднем продолжительность инкубации на 5-6 часов.
По данным Ю. М. Огороднего, резкое охлаждение зародышей ведет к падению активности каталазы и пероксидазы. Острое охлаждение зародышей с 17-го дня инкубации приводит к небольшому увеличению активности каталазы в течение первых 10 часов; после этого активность ее снижается вплоть до гибели зародыша. Резкое перегревание (до 46°) приводит к значительному нарастанию каталазы в первые 17 часов, но к моменту гибели зародыша количество ее сильно падает. Активность пероксидазы при перегреве и охлаждении поднимается, но в последнем случае более интенсивно. При высокой температуре задерживается накопление каталазы. Но с 14 по 16-й день наблюдается резкий ее рост, после чего количество ее уменьшается, но все же остается на высоком уровне. Автор утверждает, что активность пероксидазы почти не зависит от температурного фактора и чувствительность ее к нему тем меньше, чем старше зародыш. Высокая температура в инкубаторе стимулирует увеличение содержания гемоглобина, количество его резко повышается к-16-му дню и на этом уровне остается до начала вывода.
И. Я. Прицкер, определяя количество гемоглобина у цыплят через 18-24 часа после их вывода, нашел, что содержание его было тем больше, чем выше была температура воздуха во время инкубации; при температуре 38,5, 39 и 40° гемоглобина было соответственно 51,4-56%, 54,9-62,9 и 61,7-68,4%.
Как известно из работы Ю. М. Огороднего и Э. Э. Пенионжкевича, количество амниотической жидкости зависит от температуры воздуха. Повышение температуры несколько тормозит вначале накопление жидкости амниона, но последующее использование этой жидкости проходит вполне успешно. При низкой температуре жидкость амниона накапливается медленно и к 19-му дню развития зародыша в яйце находят значительное количество этой жидкости. Вязкость ее не изменяется как от высокой, так и от низкой температуры.
Можно различить несколько периодов в жизни зародыша, когда он по-разному реагирует на внешнюю температуру.
Первые 12 часов инкубации - это период, являющийся как бы продолжением развития зародыша в яйцеводе при высокой температуре. Затем до конца второго дня в случае повышенной температуры у зародыша появляются уродства вследствие нарушений роста амниона и развития центральной нервной системы. С 3 по 5-й день инкубации при повышенной температуре нарушаются отделение зародыша от желтка и формирование брюшной полости. В течение всех первых 5 дней на повышение темпе-ратуры зародыш реагирует ускорением развития и роста. С 6-го дня скорость роста под влиянием повышенной температуры начинает замедляться, и с 16-го дня у хорошо развитого зародыша появляются некоторые признаки теплокровности: при повышении температуры использование белкаи желтка уменьшается и рост зародыша задерживается, а при некотором понижении температуры использование белка и желтка усиливается и рост зародыша ускоряется.
Особое место занимает изучение колебаний температуры, периодических кратковременных изменений ее: подъем или понижение, а затем возвращение к первоначальному уровню.
В. В. Фердинандов нашел семь периодов постепенных подъемов и снижений температуры в гнездах шести наседок. Максимум температуры наблюдался на 3, 6, 10, 13, 16, 19 и 21-й день, а минимум - на 3, 5, 71/2, 101/2, 14, 17 и 20-й день. К концу насиживания амплитуда колебания температуры уменьшается последовательно по семи периодам: 7,9°, 6,8; 4,5; 6,6; 4,4; 5,0 и 3,6°. Температура также изменяется и в течение каждых суток три раза с амплитудой в 2,8°. Перемещение яиц наседкой из центра гнезда к его краям приводит к снижению температуры в среднем на 8°. Частые поворачивания яиц наседкой также обусловливают изменения температуры на верхнем уровне яиц в среднем на 2,4°. Автор зарегистрировал в среднем за 20 дней 42 перемещения яиц в сутки (35 в первые 11 дней и 48 в остальные дни).
Значение кратковременных суточных колебаний температуры В. В. Фердинандов видел в стимуляции дыхательной функции: периодические изменения внутрияйцевого давления приводят к всасыванию в яйцо кислорода и выделению газообразных продуктов обмена веществ. Однако экспериментально это интересное предположение подтверждено им не было. Он высказал также мнение, что уменьшение амплитуды колебаний температуры в последние периоды объясняется тем, что перед окончанием инкубации зародыш уже обладает собственной температурой и собственным регулирующим эту температуру аппаратом.
Во время этих исследований 30 наседок насиживали 481 яйцо и в среднем дали 85,9% вывода при колебаниях от 33,3 до 100%.
В. В. Фердинандов установил, что если тело наседки находится от яйца на расстоянии 5 см, через пять минут температура яйца снижается на 5°. Чтобы опять нагреть яйцо, требуется 50 минут. В сутки в среднем происходит 12 контактов яиц с телом птицы. Продолжительность этих контактов 60 минут каждый, а всего 12 часов. Этому соответствует количество суточных колебаний температуры внутри яйца под наседками. Колебания продолжительности обогревания отдельных яиц от 469 до 835 минут в сутки не оказывают вреда для развития зародыша. В. В. Фердинандов по этому поводу говорит, что с точки зрения потребности эмбриона такая методически колеблющаяся каждый час температура едва ли необходима. Под наседками эти колебания сведены до минимальной величины, в среднем до 1,0-1,5°. Но он полагал, что для эмбриона необходимы колебательные температуры, вызываемые биохимическими процессами в самом яйце. По мнению В. В. Фердинандова, наседка стремится усиливать подъемы температуры внутри яиц, зависящие от биохимических процессов. По мнению автора, это следует делать и в инкубаторе.
Е. Ф. Лисицким также была высказана мысль о том, что изменение температуры в довольно широких пределах, но непродолжительное, следует рассматривать как фактор, стимулирующий эмбриональный рост, как термический раздражитель. На основе этого предположения были проведены опыты (Э. Э. Пенионжкевич), в которых были получены удовлетворительные результаты инкубации.
Н. П. Третьяков также наблюдал большую изменчивость температуры яиц в гнезде наседки. По его данным, в центре гнезда температура равна 39,4°, на периферии гнезда 37,6°. Автор, не приводя экспериментальных данных, утверждает, что постоянные изменения температурыяйца в гнезде наседок создают оптимальные условия для газообмена, и считает, что колебания температуры не допускают гибели эмбрионов от перегрева.
В производственных условиях С. О. Пельтцер снижал температуру в инкубаторе "Рекорд-39" зимой до 34-35° и летом до 32-33° (по показаниям центрального термометра) два раза в сутки. На снижение и последующее восстановление температуры требовалось от 2 до 4 часов. В результате в этом инкубаторе вывод цыплят был на 1,3% выше, чем в обычных производственных партиях (Н. П. Третьяков и С. О. Пельтцер), Т. А. Залетаева инкубировала яйца в инкубаторе Рекорд-39 небольшими партиями (1-2 лотка) и выносила их из инкубатора для охлаждения на 15-20 минут два раза в день. Вывод цыплят из яиц этих лотков был на 1,5-2% выше. При этом ею было отмечено лучшее качество выведенных цыплят (Т. А. Залетаева).
Ц. X. Руус, инкубируя яйца гусей в разных инкубаторах, с 1 по 14-й день в одном из них охлаждал яйца два раза в сутки, опрыскивая водой. В другом инкубаторе яйца не охлаждали. С 15-го дня все яйца инкубировали в одном инкубаторе и два раза в сутки охлаждали по 30-50 минут, а затем опрыскивали водой. Автор пришел к выводу, что опрыскивание гусиных яиц водой в первую половину инкубации тормозит рост зародыша, но вместе с тем способствует лучшему развитию сердца, более высокому содержанию гемоглобина в крови во вторую половину инкубации и лучшему усвоению питательных веществ. К сожалению, в работе не приведены данные о конечных результатах инкубации.
Е. Ф. Лисицкий, А. У. Быховец и Г. С. Крок пришли к заключению, что периодические повышения (раз в 3 или 4 дня до 40°) температуры до 19-го дня инкубации и последующие снижения (охлаждения вне инкубатора) с 17 по 24-й день два раза и с 25 по 28 день три раза в сутки до 35° благоприятно влияют на эмбриональное развитие гусей.
Однако целесообразность применения колебательных температур требует еще серьезного теоретического обоснования и практической проверки. Этот прием не нашел применения в практике инкубации.
Имеющиеся данные указывают лишь на способность зародышей птицы переносить иногда довольно значительные отклонения температуры от среднего уровня.
Способность зародышей переносить значительные изменения температуры используется при инкубации яиц уток и гусей в инкубаторах, не обеспечивающих удаления избытков тепла яиц во вторую половину инкубации. В таких случаях прибегают к различным приемам охлаждений: выносят яйца из инкубатора, периодически значительно снижают температуру воздуха инкубатора, опрыскивают яйца водой. Эти приемы позволяют получить хорошие результаты инкубации.
То обстоятельство, что при создании в инкубаторе условий для удаления излишков тепла (снижение температуры воздуха инкубатора, увеличение скорости движения воздуха около яиц, увеличение обмена воздуха в инкубаторе и т. п.) получаются хорошие результаты без охлаждения яиц, ставит под сомнение биологическую целесообразность неоднократных ежедневных охлаждений яиц уток и гусей.
Однако эти соображения не исключают возможности благоприятного воздействия на организм зародыша значительных периодических снижений температуры и последующего ее восстановления. Повышение-жизненности зародыша таким воздействием было показано на самых ранних стадиях развития: после нагревания и последующего охлаждения яиц в первые 12-15 часов инкубации срок хранения яиц, после которого зародыши развиваются нормально, увеличивался до 20-25 дней(против 3-5 дней), а вывод цыплят, гусят, утят и индюшат повышался по сравнению с выводом из свежих яиц, не подвергавшихся такому воздействию. Есть все основания полагать, что и в последующее время зародышевого развития возможен такой же эффект от воздействия пониженной температурой.
Из данных В. В. Хаскина, полученных при изучении в отдельные дни инкубации колебаний температуры воздуха от 30 до 42° и обратно в течение 1,5-2 часов, следует, что в последнюю четверть периода инкубации яиц уток такие изменения повышают газообмен на 25-35 %.
Н. П. Третьяков пришел к выводу, что охлаждение яиц кур и уток в течение часа при температуре 16-18-24° на любой стадии развития эмбрионов не вызывает снижения вывода и ухудшения качества птенцов. Г. А, Машталлер, подвергая в своих опытах яйца кур, уток и гусей одно- и двукратному за период инкубации охлаждению в течение 40-50, а иногда 60 минут при температуре 1-5°, получал увеличение вывода молодняка до 20%. Этот прием повышения биологической ценности инкубируемых яиц заслуживает серьезного изучения с целью практического его применения.
Диагностика ошибок режима инкубирования (часть третья)
Диагностика недогрева. Низкая температура задерживает развитие зародыша с первых дней инкубации, но не вызывает таких глубоких и специфических нарушений в развитии, как высокая температура. При просвечивании яиц после 6 дней инкубации обнаруживается общее отставание развития: зародыши малы, лежат близко к скорлупе, благодаря чему ясно различимы, кровеносная система на желтке развита слабо, кровеносные сосуды слабо наполнены кровью и имеют прозрачный розовый цвет, зародыши мало подвижны.
Зародыши погибают поздно. Кровяные кольца малы и бледны. Вскрытие яиц показывает недоразвитие оболочек и анемию зародышей (Г. К. Отрыганьев и Г. И. Крылов).
Рост аллантоиса при недогреве сильно задерживается и замыкание его краев происходит с большим опозданием. Поэтому при просвечивании яиц после 11 дней инкубации обнаруживается, что аллантоис замкнут менее чем у 50% яиц.
И. Я. Прицкер указывает, что при недогреве пух у цыплят развивается хуже, чем при нормальной температуре инкубации или при перегреве. Во время просвечивания перед выводом также видно общее отставание в развитии зародыша: он мал, не заполняет яйцо, которое просвечивается как в остром конце, так и у воздушной камеры; последняя имеет небольшие размеры; выпячивание шеи в воздушную камеру происходит с большим опозданием.
Наклев скорлупы начинается недружно и тоже с большим опозданием, но в надлежащем месте, и скорлупа отламывается крупными кусками.
Вывод идет недружно и продолжается очень долго, иногда несколько дней. Выведенный молодняк хорошо опушен. Пупочное кольцо хорошо заживлено и не имеет никаких рубцов. Остаточный желток в большинстве случаев невелик. Выведенный молодняк очень мало подвижен, вял, плохо и неуверенно стоит на ногах. Скорлупа, оставшаяся после вывода, имеет бледно-розовый или светло-кремовый цвет благодаря слабому наполнению кровью сосудов аллантоиса (И. Я. Прицкер).
При очень сильном и длительном недогреве выведенные цыплята имеют большой остаточный желточный мешок, часто страдают поносом, оставшаяся после вывода скорлупа грязная, сырая, с неиспользованным белком (Г. К. Отрыганьев).
концу вывода остается много яиц с наклевом и живыми зародышами, которые слабы и не могут разломать скорлупу, чтобы от нее освободиться. Попытки оказать им помощь при выводе приводят к кровоизлияниям из сосудов аллантоиса и к гибели зародышей.
При вскрытии яиц с задохликами оказывается, что много живых зародышей имеется и в яйцах без наклева. Большинство зародышей оформившихся, с втянутыми желтками и использованным белком; на голове и шее у них наблюдается отек больших размеров, часто гипереми-рованный и с кровоизлияниями.
Только при очень сильном недогреве желток остается невтянутым и белок неиспользованным. Желточный мешок при недогреве бледный, пупочное кольцо не замкнуто, а белок чаще всего мутный, жидкой консистенции. Очень часто весь желток или отдельные его участки ярко-зеленого цвета.
При вскрытии задохликов можно наблюдать анемию внутренних покровов и органов. Кишечник переполнен желтком и каловыми массами, особенно прямая кишка, диаметр которой иногда достигает толщины с палец, печень увеличена (Г. К. Отрыганьев). Сердце увеличено (Е. Ф. Лисицкий) и анемично (И. Я. Прицкер) (рис. 3,а).

Рис. 3. а - вскрытый задохлик при недогреве, б - проклев скорлупы при высокой влажности, в - проклев скорлупы при низкой влажности, г - вскрытый задохлик при высокой влажности
Нарушение влажности. Относительная влажность воздуха инкубатора оказывает существенное влияние на обмен веществ и развитие зародышей. Она регулирует испарение воды из яиц в течение большего периода инкубации и регулирует теплоотдачу.
Влажность в отличие от температуры, влияние которой сказывается почти одновременно с началом воздействия, оказывает действие более медленно и для своего появления требует известного времени. Однако, постепенно накапливаясь, неблагоприятное воздействие отклонений влажности от нормы очень велико и не всегда поправимо.
Диагностика высокой влажности. В первые дни инкубации высокая влажность отрицательного влияния на развитие зародыша не оказывает.
При инкубации грязных яиц при высокой влажности как в первые дни, так и в течение всего периода инкубации могут развиваться в яйцах гнилостные процессы и появляться "тумаки".
Высокая влажность после 6-го дня инкубации начинает задерживать развитие, в связи с чем замыкание краев аллантоиса запаздывает.
Некоторое повышение смертности зародышей под влиянием высокой влажности в средние дни инкубации не сопровождается какими-либоспецифическими признаками нарушений. Наблюдается общее отставание в росте и развитии зародышей и их оболочек. Характерна малая величина воздушной камеры вследствие недостаточной потери веса яйцами (менее 0,7-0,6% в сутки).
Внешний вид яиц при просвечивании перед выводом очень напоминает вид яиц, инкубируемых при недогреве. Воздушная камера очень мала; выпячивание шеи зародышем не начинается. Большие просветы в остром конце яйца и у воздушной камеры указывают на значительные количества околоплодных жидкостей.
Начало наклева задерживается (до 21 дня) и проходит недружно. Подскорлупная оболочка после проклева буреет, и часто после этого дальнейший проклев прекращается вследствие гибели зародыша.
По данным Э. Э. Пенионжкевича и Н. М. Шкляра, высокая влажность вызывает характерную форму "наклева с выделением жидкости". Эта жидкость быстро засыхает и закрывает отверстие в скорлупе и зародыш погибает. Жидкостью может приклеиться клюв зародыша к скорлупе, что приведет к прекращению движения зародыша и его гибели (Г. К. Отры-ганьев). Попытки оказать помощь при выводе обычно вызывают кровотечение и смерть зародыша (рис. 3, б). У выведенного молодняка пух, особенно у пуповины и анального отверстия, обычно грязный. Пигментация конечностей и пуха очень слабая. Цыплята вялые, мало подвижные; живот у них очень большой, но может быть и мягкий, вследствие втягивания большого жидкого желтка.
Смерть большинства невылупившихся цыплят наступает в момент проклева, от захлебывания околоплодной жидкостью. При вскрытии задохликов характерно обилие клейкой слизи в плодовых оболочках, переполнение жидкостью кишечного тракта, легкие гиперемированы, воздуха не содержат.
У мертвых зародышей наблюдается отек шеи и головы, большой вздутый зоб, наполненный жидкостью.
Другие поражения внутренних органов такие же, как и при недогреве.
Диагностика пониженной влажности. Очень низкая влажность воздуха в первые дни инкубации вызывает некоторое повышение смертности зародышей, но не ведет к появлению каких-либо специфических поражений их. Пониженная влажность усиливает проявление признаков перегрева при повышенной температуре (Г. К. Отрыганьев и Е. Н. Кучковская).
Яйцо очень теряет в весе (более 0,5-0,6%), и воздушная камера быстро увеличивается в объеме. Аллантоис может замыкаться раньше срока.
Во время просвечивания яиц перед выводом обнаруживается несколько ускоренное развитие многих зародышей, мертвых зародышей мало.
Наклев и вывод начинаются раньше срока. Подскорлупные оболочки очень сухи и прочны. Зародыш, пробивая скорлупу, не в состоянии разорвать подскорлупные оболочки, от которых отваливаются кусочки скорлупы. Он может совершить полное круговое движение и не освободиться от скорлупы.
После проклева скорлупы пух очень быстро высыхает. Даже небольшой участок высохшего в скорлупе пуха на корпусе зародыша мешает его движениям, а иногда движения прекращаются и зародыш погибает.
Вывод затруднен и протекает медленно. Выведенные цыплята мелкие, плохо опушенные, но подвижные. Пух их интенсивно пигментирован.При вскрытии яиц с задохликами можно обнаружить признаки, характерные для перегрева, но в ослабленной форме.
По данным Г. К. Отрыганьева, у задохликов, как правило, бывают кровоизлияния в аллантоисе за счет ранения клювом еще функционирующих кровеносных сосудов; возле клюва большой сгусток крови.
Диагностика недостаточной вентиляции. Загрязнение воздуха инкубатора оказывает отрицательное влияние на развитие зародышей, но специфических диагностических признаков плохой вентиляции пока не найдено.
По Г. К. Отрыганьеву, газообмен яйца может быть нарушен вследствие недостаточной вентиляции, при закупорке пор скорлупы грязью, содержимым соседних разбитых яиц. При нарушении газообмена рост и развитие задерживаются. В средние дни инкубации резкие удушья (асфиксии) вызывают те же ненормальности в развитии, что и острый перегрев: переполнение сосудов аллантоиса кровью, гиперемию, кровоизлияния в кожу. Характерный признак - наличие крови в амниоти-ческой жидкости (гематоамнион). Недостаточный газообмен в период инкубации вызывает неправильные положения эмбриона в яйце, поэтому проклев скорлупы происходит в остром конце яйца.
Нарушения, связанные с положением и поворачиванием яиц. Отсутствие поворачивания яиц приводит к большому количеству прилипаний и присыханий оболочек и зародышей к скорлупе. В отличие от присыха-ний, появляющихся при высокой температуре и низкой влажности, при отсутствии поворачивания яиц все мертвые и присохшие зародыши лежат с одной стороны (верхней).
Если яйца не поворачивают, то наблюдается повышение смертности зародышей при выводе.
Недостаточное количество поворачиваний или недостаточный угол поворота яиц вызывают прежде всего слабое и неправильное развитие аллантоиса, который не замыкается в остром конце яйца или замыкается очень поздно. При этом встречается много яиц, в которых аллантоис растет своими краями над белком и, замыкаясь, оставляет вне себя белок в остром конце яйца. Молодняк в таком случае выводится мелкий и слабый.
Нарушения, связанные со скоростью движения воздуха. Скорость движения воздуха не оказывает влияния непосредственно на развивающиеся зародыши, но усиливает или ослабляет влияние других внешних факторов - температуры, влажности.
В шкафных инкубаторах скорость движения воздуха обеспечивает однородность режима во всех его точках.
Неравномерное развитие зародышей во всех местах инкубатора и неодновременное начало вывода косвенно указывают на наличие зон, в частности температурных, вследствие недостаточной скорости движения воздуха.
Прижизненная оценка развития зародышей (часть третья)
По данным Н. П. Третьякова, из яиц, имеющих ломаную очерченность границ пуги, и из яиц, имеющих ровную очерченность пуги,- получали разный вывод - 80,5 и 56,2, 89,0 и 32,0% от числа яиц уток, перенесенных на вывод, и 96,1 и 70,9 из яиц кур.Разница в выводе из яиц кур с хорошо развитыми и с отсталыми зародышами на 19-й день инкубации также очень существенна, что видно из таблицы 13, составленной по данным М. В. Орлова и Е. Н. Кучковской.
Таблица 13. Вывод цыплят из яиц с хорошо развитыми и отсталыми зародышами к 19-му дню инкубации (в %)
|
Порода кур |
Яйца с хорошо развитыми зародышами |
Яйца с отсталым развитием зародышей |
Разница в проценте вывода |
|
Леггорн (белые) |
97,2 |
71,6 |
25,6 |
|
Нью-гемпшир |
91,7 |
78,1 |
13,6 |
|
Род-айланд |
98,2 |
66,7 |
31,5 |
Следует отметить, что выпячивание шеи зародышем в воздушную камеру не является признаком безусловно хорошего развития, если острый конец яйца не заполнен телом зародыша.
Важный признак хорошего развития зародышей - продолжительность инкубационного периода. Если зародыш хорошо питается и развивается, то инкубационный период его заканчивается своевременно. Если развитие зародыша и его обмен веществ нарушаются либо под влиянием неполноценности яйца, либо под влиянием несоответствия режима инкубирования требованиям зародыша, то в большинстве случаев это ведет к удлинению инкубационного периода. В таком случае вывод молодняка начинается позже и продолжается дольше.
При хорошем развитии наклев скорлупы зародышем начинается своевременно и проходит дружно. Наклев бывает ближе к тупому концу яйца, и скорлупа, высоко поднимаясь, отламывается крупными кусками. После проклева зародыш энергично совершает круговое движение внутри яйца. Подскорлупная оболочка эластична, она разрывается по мере разрушения скорлупы и естественный белый цвет ее сохраняется.
По данным И. Я. Прицкера, средняя продолжительность процесса освобождения цыпленка из скорлупы с момента наклева 9,8- 12,5 часа.
Вывод молодняка при хорошем развитии зародышей проходит в следующие сроки (табл. 14).
Таблица 14. Вывод молодняка при хорошем развитии зародыша
|
Молодняк |
Начало вывода |
Массовый вывод |
Конец вывода |
|
Цыплята |
В конце 20 дня |
Первая половина 21 дня |
В конце 21 дня |
|
Утята и индюшата |
26 день |
27 день |
В конце 27 или в начале 28 |
|
Гусята |
29 день |
30 день |
В конце 30 или в начале 31 |
Нарушение развития зародышей в отдельных случаях может и не привести к значительному повышению их гибели, и хотя процент вывода может быть высоким, но полученный молодняк будет ослабленным, а продолжительность инкубации, как правило, увеличивается. Необходимостремиться к тому, чтобы при просмотре не менее 75-80% яиц было с хорошо развитыми зародышами и чтобы вывод протекал своевременно. Важно не только получить при выводе много молодняка, но получить его во-время. Это будет характеризовать хорошее его качество.
Вывод цыплят из яиц с признаками хорошего развития зародышей бывает более высоким и своевременным, чем из яиц с признаками отсталого развития зародышей. Цыплята в первом случае получаются более крупными и вес остаточного желтка меньше. Кроме того, молодки, хорошо развившиеся в эмбриональный период, более жизнеспособны, достигают половой зрелости при большом живом весе, имеют более раннюю скороспелость и лучшую яйценоскость, особенно осенью и зимой.
Вес выведенного и обсохшего молодняка составляет около 65% от первоначального веса яиц. У цыплят остаточный желток мал и живот хорошо подобран. Пупочное кольцо хорошо замкнуто и не имеет никакого шрама. У белоногих пород плюсны и клюв розовато-желтые или оранжевые. Молодняк крепко стоит на ногах, подвижен.
В практической работе не вскрывают яйца с живыми зародышами для характеристики их развития. Однако целесообразно на ранних стадиях инкубации проводить пробные вскрытия из каждой партии однородных яиц (по 10 штук).
Г. К. Отрыганьев рекомендует такое вскрытие делать через 36 часов после начала инкубации. Хорошее развитие зародыша в это время характеризуется следующими показателями: диаметр сосудистого поля 6-8 мм, длина зародыша 4-5 мм, число сомитов 8-10 пар.
По материалам Н. А. Коноплева, к 48 часам насиживания и в инкубаторах при хорошем развитии зародыша сосудистое поле имеет диаметр не менее 12 мм, длина зародыша не менее 7 мм, сомитов не менее 24 пар.
Прижизненная оценка развития зародышей (часть четвертая)
Зародыш должен занимать правильное положение: вдоль малой оси яйца головой вправо, если яйцо расположено тупым концом от наблюдателя, а острым к наблюдателю. Если зародыш занимает неправильное положение, то это указывает на неудовлетворительное его развитие.
В таблице 15 приведены данные Г. К. Отрыганьева по весу зародышей кур породы белый леггорн и пекинских уток. Вывод из яиц кур в инкубаторе "Рекорд-39" был 85% а из яиц уток-81%.
Таблица 15. Вес зародышей кур и уток (в г)
|
Дни инкубации |
Куры |
Утки |
|
3,5 |
0,05-0,06 |
-- |
|
4,5 |
0,13-0,19 |
-- |
|
5,5 |
0,18-0,40 |
0,14-0,15 |
|
6,5 |
0,42-0,90 |
0,25-0,37 |
|
7,5 |
0,75-1,25 |
0,53-0,60 |
|
8,5 |
0,80-2,10 |
0,65-0,89 |
|
9,5 |
1,70-2,60 |
1,20-1,45 |
|
10,5 |
2,70-3,50 |
1,90-2,00 |
|
11,5 |
3,65-4,95 |
2,55-2,75 |
|
12,5 |
5,30-6,80 |
2,75-3,90 |
|
13,5 |
7,30-9,70 |
4,55-5,95 |
|
14,5 |
10,30-11,70 |
7,00-8,00 |
|
15,5 |
12,40-17,00 |
10,00-11,30 |
|
16,5 |
16,55-20,20 |
11,75-14,25 |
|
17,5 |
19,30-24,80 |
15,10-18,30 |
|
18,5 |
23,10-29,10 |
18,60-23,00 |
|
19,5 |
28,20-35,40 |
24,30-27,70 |
|
20,5 |
-- |
26,60-32,40 |
|
21,5 |
-- |
24,85-35,90 |
|
22,5 |
-- |
40,60-41,60 |
|
23,5 |
-- |
42,50-45,50 |
|
24,5 |
-- |
43,80-46,95 |
|
25,5 |
-- |
44,10-47,10 |
К приемам прижизненного контроля относится также и учет потери веса яйцами во время инкубации. Испарение воды из яйца происходит и под влиянием внешних условий (особенно в первую половину инкубации) и под влиянием интенсивности обмена веществ (особенно во вторую половину инкубации). Потеря в весе яиц как результат испарения воды и обмена газов по периодам инкубации отражает интенсивность обмена веществ зародыша, связанную с качеством яиц и условиями инкубации.
Среднесуточная потеря в весе яиц в начале инкубации при удовлетворительных результатах инкубации бывает обычно в следующих пределах: для яиц кур с 1 по 6 день - 0,5-0,6%, для яиц уток с 1 по 7 день - 0,4-0,5% и для яиц гусей с 1 по 8 день инкубации - 0,3-0,4%.
После того как аллантоис покроет все содержимое яйца, среднесуточная потеря веса значительно возрастает, и тем больше, чем интенсивнее идет развитие зародыша. У яиц кур она составляет более 0,6% (0,7-0,8%). В отдельные сутки потеря веса может доходить до одного и более процентов. Яйца уток теряют 0,5% и более, а яйца гусей 0,4% своего первоначального веса и более в среднем за сутки.
P.S: Авторские права статьи не известны. Поэтому, если Вы являетесь автором и можете это подтвердить, смело обращайтесь к администрации сайта для урегулирования данного вопроса!